Objective 1. Compare understory plants communites of treatment types for significant differences
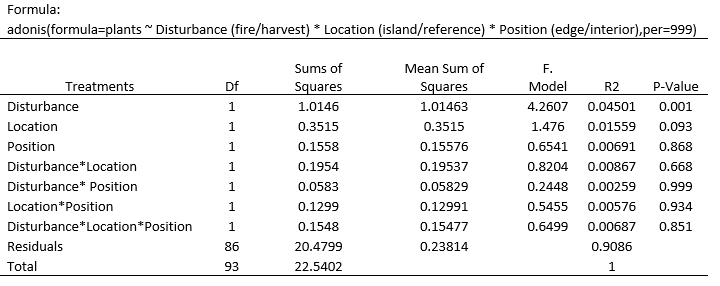
Table 3. Results of a permutational multivariate analysis of variance (PerMANOVA) of a Bray-Curtis distance matrix of the understory plant community
|

Figure 19. Non-metric Multidimensional Scaling plot (NMDS) of understory plant abundance, using Bray-Curtis data with a confidence interval of 0.8.
|

Figure 20. Figure 19. Non-metric Multidimensional Scaling plot (NMDS) of understory plant abundance, using Bray-Curtis data with a confidence interval of 0.8. Species vectors are presented in black. Edge plot confidence elipses represented by dashed lines
|
According to the results of a permutational multivariate analysis of variance (Table 3), there is a significant difference between the communities found in the fire plots and those found in the harvest plots. There was no statistical difference found between the island remanant and reference communites within the harvest and fire sites, and no statistical difference found between the edge and interior communites within the harvest and fire sites. As seen in the NMDS plots, there is significant overlap between the communities of both edge/interior plots, and the island/reference plots.
|
Species Associated with Fire Plots
ARANUD |
ABIBAL |
GYMDRY |
LYCANN |
VACMYR |
LINBOR |

Picture 1. Wild sarsparilla (Aralia nudicaulis). Image courtesy of Go Botany
|

Picture 2. Balsam fir (Abies balsamea). Image courtesy of Go Botany
|

Picture 3. Oak fern (Gymnocarpum dryopteris). Image courtesy of Go Botany
|

Picture 4. Clubmoss (Lycopodium annotium). Image courtesy of Bernd Haynold
|

Picture 5. Blueberry (Vaccinium myrtilloides). Image courtesy of the Agroforestry & Woodlot Extension Society
|

Picture 6. Twinflower (Linnaea borealis). Image courtesy of Go Botany
|
Species Associated with Harvest Plots
CALCAN |
RUBPUB |
RIBTRI |
ROSACI |
GALBOR |
FRAVIR |

Picture 7. Bluejoint grass (Calamagrostis canadensis). Image courtesy of Rob Routledge
|

Picture 8. Dewberry (Rubus pubescens). Image courtesy of Albert Herring
|

Picture 9. Red currants (Ribes triste). Image courtesy of Melody Lytle
|

Picture 10. Prickly rose (Rosa acicularis). Image courtesy of sydcannings
|

Picture 11. Northern bedstraw (Galium boreale). Image courtesy of Barry Breckling
|

Picture 12. Strawberry (Fragaria virginiana). Image courtesy of Seven Oaks Nursery
|
Overall, there is a large amount of overlap between the understory communities of fire and harvest plots. There are however 2 key differences: First, there is a greater dispersion among fire plots than harvest plots, indicating a greater diversity of species (Figure 18). Second, the species most often associated with harvest plots are extremely common species that are usually associated with deciduous dominated medium nutrient mesic ecosites (our target ecosite), while plants associated with fire plots are rarer in those ecosites (Figure 19) (Beckingham & Archibald, 1996). I had hypothesized that differences between fire and harvest plots would primarily occur in the edges, where harvest plots would be more succeptible to edge effects than fire sites. Based on the results of the PerMANOVA presented in table 3, both fire and harvest edges have similar understory communities to interior forest, thus its unlikely that any difference between harvest and fire plots is due to diverging edge types.
Rather, I believe the difference between fire and harvest plots has more to do with how the disturbances create the remnants. The process of creating harvest remnants begins with cutblock selections. Planned harvests happen exclusively within areas containing medium-high tree productivity and lots of merchantable timber. Cutblocks are fairly homogenous units, both in terms of topography, forest composition, nutrient regime, and moisture. Additionally, legal requirements and best management practices stipulate that harvests must not be done too closer to waterways, and patch remnants must be representative of the forest that was harvested. Our choice to selct deciduous dominated forest with medium nutrient regimes and mesic moisture was intentional, as this forest type is commonly harvested in the boreal. This resulted in remants that conformed very readily with our desired site types- they had already been selected for harvest by the forestry companies based on similar criteria.
Fire remnants are different- they are created by fire behaviour and landscape features. Topographical features like hills, valleys, and ravines and hydrological features like lakes, wetlands, and streams all impede fires, and thus fire remants are more likely to be associated with these topographical features. Additionally, fire remants can be created when a burn within one forest type encounters a less flammable forest type. For example, pines and spruce are more flammable than aspen, so a fire that burns through a coniferous forest may stop when it encounters a patch of aspen, creating a fire remant. This variability also means we had difficulty contolling the ecosite types of our fire remnants, as we were limited to what resulted from the natural processes.
Effectively, this means these remnants are created through quite different processes. Harvest remnants are placed in a relatively topographically homogenous place away from hydrological features, and they are chosen to be representative of the forest that is harvested. Fire remnants are the oposite in the sense that they are often located in more heterogenous landscapes, near hydrological feature, and are oftetimes created expressely because they contain different vegetation than the burned area. I believe that the different ways in which these remnants form is leading to the difference in understory species. For example, both the utikuma complex fire and the M024 fire burned within areas dominated by black spruce/pine forest. I believe that the remants found in those fires were created when the fire stopped burning when it collided with patches of aspen, burning only the surrounding coniferous forest. When two different ecosite types border each other, they tend to "share" understory species- that is to say species that are common to one ecosite will appear in the other and vice versa. One thing I notice about Utikuma and M024, is their associations with Labrador tea (LEDGRO), blueberry (Picture 5, VACMYR), and bog cranberry (VACVIT), species that are much more common in poorer nutrient ecosites like black spruce/pine dominated forest. These species would appear less often in harvest remants, beacuse they dont border different ecosites in which those plants are common.
The flattop fire was located in a hilly area, and thus our remnants were often located on the tops of hill, within ravines, or adjacent to creeks. These plots were associated with fern species such as oak fern (Picture 3. GYMDRY), ostrich fern (MATSTR), clubmoss (Picture 4, LYCANN), and lady fern (ATHFIL), as well as balsam fir Picture 2, ABIBAL) and starflower (TRIBOR). All of these plants are shade tolearant plants that thrive in cool, moist ecosites (like ravines and creeks), and are often characteristic of late seral forest (Abrahamson, 2022), indicating that these areas could be a persistent fire refugia. And again, these species would not be present within are harvest sites because those are placed in a different ecosite type on relatively flat ground away from hydrological features.
Rather, I believe the difference between fire and harvest plots has more to do with how the disturbances create the remnants. The process of creating harvest remnants begins with cutblock selections. Planned harvests happen exclusively within areas containing medium-high tree productivity and lots of merchantable timber. Cutblocks are fairly homogenous units, both in terms of topography, forest composition, nutrient regime, and moisture. Additionally, legal requirements and best management practices stipulate that harvests must not be done too closer to waterways, and patch remnants must be representative of the forest that was harvested. Our choice to selct deciduous dominated forest with medium nutrient regimes and mesic moisture was intentional, as this forest type is commonly harvested in the boreal. This resulted in remants that conformed very readily with our desired site types- they had already been selected for harvest by the forestry companies based on similar criteria.
Fire remnants are different- they are created by fire behaviour and landscape features. Topographical features like hills, valleys, and ravines and hydrological features like lakes, wetlands, and streams all impede fires, and thus fire remants are more likely to be associated with these topographical features. Additionally, fire remants can be created when a burn within one forest type encounters a less flammable forest type. For example, pines and spruce are more flammable than aspen, so a fire that burns through a coniferous forest may stop when it encounters a patch of aspen, creating a fire remant. This variability also means we had difficulty contolling the ecosite types of our fire remnants, as we were limited to what resulted from the natural processes.
Effectively, this means these remnants are created through quite different processes. Harvest remnants are placed in a relatively topographically homogenous place away from hydrological features, and they are chosen to be representative of the forest that is harvested. Fire remnants are the oposite in the sense that they are often located in more heterogenous landscapes, near hydrological feature, and are oftetimes created expressely because they contain different vegetation than the burned area. I believe that the different ways in which these remnants form is leading to the difference in understory species. For example, both the utikuma complex fire and the M024 fire burned within areas dominated by black spruce/pine forest. I believe that the remants found in those fires were created when the fire stopped burning when it collided with patches of aspen, burning only the surrounding coniferous forest. When two different ecosite types border each other, they tend to "share" understory species- that is to say species that are common to one ecosite will appear in the other and vice versa. One thing I notice about Utikuma and M024, is their associations with Labrador tea (LEDGRO), blueberry (Picture 5, VACMYR), and bog cranberry (VACVIT), species that are much more common in poorer nutrient ecosites like black spruce/pine dominated forest. These species would appear less often in harvest remants, beacuse they dont border different ecosites in which those plants are common.
The flattop fire was located in a hilly area, and thus our remnants were often located on the tops of hill, within ravines, or adjacent to creeks. These plots were associated with fern species such as oak fern (Picture 3. GYMDRY), ostrich fern (MATSTR), clubmoss (Picture 4, LYCANN), and lady fern (ATHFIL), as well as balsam fir Picture 2, ABIBAL) and starflower (TRIBOR). All of these plants are shade tolearant plants that thrive in cool, moist ecosites (like ravines and creeks), and are often characteristic of late seral forest (Abrahamson, 2022), indicating that these areas could be a persistent fire refugia. And again, these species would not be present within are harvest sites because those are placed in a different ecosite type on relatively flat ground away from hydrological features.
Objective 2: Examine the role of forest structure on understory community assemblages and microclimatic conditions

Figure 21. Indirect gradient principle component analysis of modified forest structure data. Displaying principle components 1 & 2, as well as their species association vectors
As mentioned before, PC1 is a gradient of tree cover and tree mortality. The negative side of the PC1 axis is plots with high canopy covers, and high amounts of living tree basal areas. There were no strong understory species associations with these plots, save a minor association with kidney-leaved violet (VIOREN). An example of such a site is presented in picture 13, note the large amount of large (for the region) living trees, and absence of dead trees and coarse woody debris. The positive side of the PC1 axis contains plots with a lot of dead trees and coarse woody debris. These plots were associated with disturbance adapted species like fireweed (CHAANG), and light-loving shrubs like bracted honeysuckle (LONINV), lowbush cranberry (VIBEDU), and red osier dogwood (CORSTO). An example of such a site is presented in picture 14, note the open canopy, abundant deadwood and developped shrub layer. The positive side of PC2 is mostly comprised of plots with highsapling counts and/or high basal areas of living deciduous trees. These plots were associated with green alder (ALNCRI), and wild sarsparilla (ARANUD). An example of such a site is presented in picture 16, note the open abundand amount of saplings and shrubs. The negative side of PC2 is plots with high coniferous tree basal areas and high microclimatic temperatures. There are two categories of understory species associated with these plots: shrubs like willows (SALIX) and buffaloberry (SHECAN), and forbs like horsetail (EQUPRA), wintergreen (PYRASA), and twinflower (LINBOR). An example of such a site is presented in picture 15, not the spruce trees, and horsetail abundant understory.

Picture 13. Harvest reference interior plot ALP-13-RI. Representative of negative PC1 plot (Figure 21 -1.6, 0.27)
|

Picture 14. Harvest island edge plot MEE-15-IE. Representative of positive PC1 plot (Figure 21 6.6, 2.5)
|

Picture 15. Fire island edge plot M024-07-IE. Representative of negative PC2 plot (Figure 21 -0.61, -2.28)
|

Picture 16. Harvest island island plot ALP-08-II. Representative of positive PC2 plot (Figure 21 -2.52, 3.02)
|
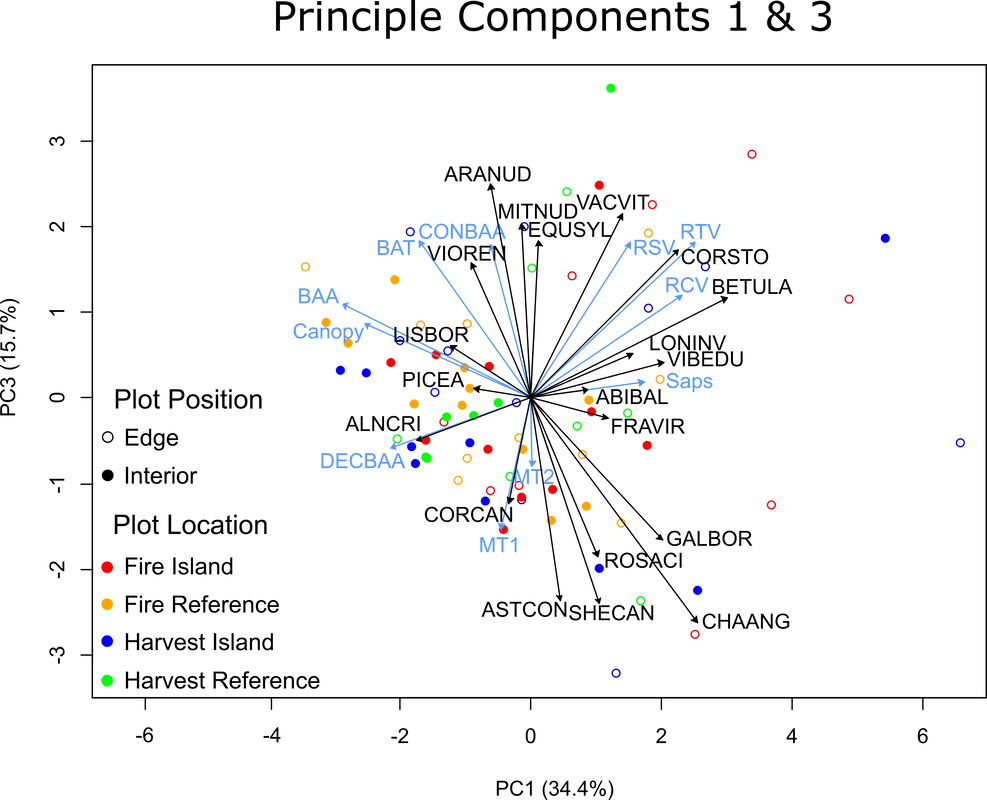
Figure 22. Indirect gradient principle component analysis of modified forest structure data. Displaying principle components 1 & 3, as well as their species association vectors
The negative side of PC3 is comprised of plots with higher microclimatic temperatures, low canopy cover and few coniferous trees. Species associated with the plots include buffaloberry (SHECAN), bunchberry (CORCAN), showy aster (ASTCON), fireweed (CHAANG), and rose (ROSACI). An example of such a plot is presented in picture 17, note the open canopy, lack of conifers and abundant understory cover. The positive side of PC3 is generally associated with high basal areas of living conifer trees, but slightly diverges based on its association with PC1. Some sites have higher canopy covers and are associated with small shade torelant forbs like the kidney-leaved violet (VIOREN), bishops cap (MITNUD), and forest horsetail (EQUSYL). An example of such a plot is presented in picture 18, note the large living conifer trees and low amount of understory cover. Other sites have higher volumes of dead trees, and are asociated with shrubs like red osier dogwood (CORSTO). An example of such a plot is presented in picture 19, note the large conifer trees, abundant shrub layer, and deadwood.

Picture 17. Harvest island interior plot MEE-13-II. Representative of negative PC3 plot (Figure 22, 2.56, -2.25)
|

Picture 18. Fire reference interior plot S057-05-RI. Representative of positive PC3 plot (Figure 22, -2.08, 1.38)
|

Picture 19. Harvest reference edge plot ALP-08-RE. Representative of positive PC3 plot (Figure 22, 0.03, 1.53)
|
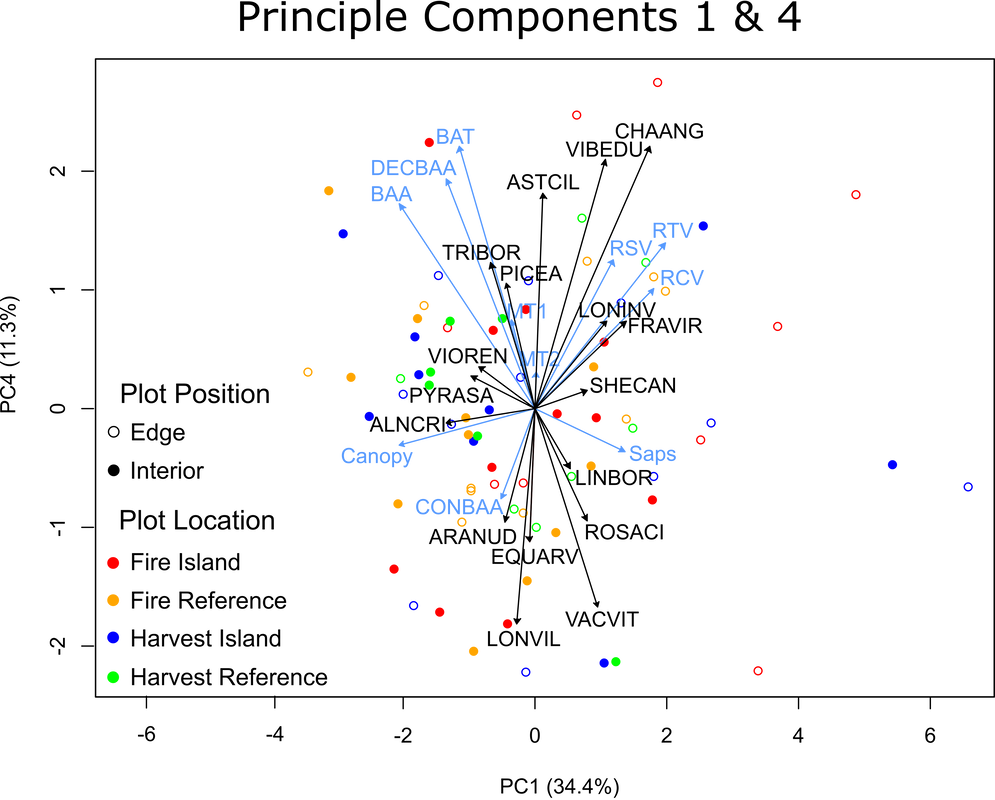
Figure 23. Indirect gradient principle component analysis of modified forest structure data. Displaying principle components 1 & 4, as well as their species association vectors
The negative side of PC4 is comprised of plots with high basal areas of conifer trees, but are relatively low in total tree basal area and little to no deadwood. these plots are associated with wild sarsparilla (ARANUD), horsetail (EQUARV), fly honeysuckle (LONVIL), and bog cranberry (VACVIT). An example of such a plot is presented in picture 20, note the large amount of small DBH conifer trees and forb understory. The positive side of PC4 is comprised of plots with slightly higher temperatures and high total and deciduous basal areas, while also associating with higher volumes of deadwood. Species associated with higher basal areas of living trees include the shade tolerant starflower (TRIBOR), and spruce seedlings (PICEA), while species associated with more deadwood include the shade intolerant fireweed (CHAANG), bracted honeysuckle (LONINV), and strawberries (FRAVIR). Examples of such plots are presented in pictures 21 & 22. Note the abundant deadwood in picture 21, and the abundance of deciduous trees in picture 22.

Picture 20. Fire reference interior plot S057-11-RI. Representative of negative PC4 plot (Figure 23, -0.93, -2.05)
|

Picture 21. Fire island edge plot S056-05-IE. Representative of positive PC4 plot (Figure 23, 1.87, 2.74)
|

Picture 22. Fire island interior plot M024-07-II. Representative of Positive PC4 plot (Figure 23, -1.60, 2.24)
|
Using the information derived from the above summaries of different forest structures found in my experiment we can answer our research questions:
First of all, we can see that temperature has an orthogonal or slightly oppositely coorelated association with microclimate temperatures, indicating canopy cover has little to no effect on microclimate. Its a similar story with tree mortality: Temperatures are slightly corelated in figure 21, slightly negatively correlated in figure 22, and are correlated in figure 23, but the small length of the temperature vector indicates minor importance. Overall, I cant conclude that either tree canopies or recent deadwood has any significant effect on microclimatic hypothesis. My thought is that high understory covers in plots with high tree mortality are shading the microclimate sensors, resulting in lower temperatures for plots with less shading from the canopy.
Second, we can see that higher tree basal areas and canopy covers are associated with understory species that are shade tolerant, and late succesional. The association of species can depend on the type of canopy, with certain species like bog cranberry (VACVIT) and bishop's cap (MITNUD) preferring conifer forests, while a species like bunchberry (CORCAN) preferred deciduous canopies. Other species like wild sarsparilla (Picture 1, ARANUD), kidney-leaved violet (VIOREN), and wintergreen (PYRASA) occured readily under both canopy types. All the aforementioned species are shade tolerant, mid to late successional species that would decrease in importance after disturbance (Abrahamson, 2022).
Finally, we can also see that certain plants favour sites with higher volumes of recent deadwood: I found shade intolerant early succession plants such as fireweed (CHAANG), strawberry (FRAVIR), and birch seedlings (BETULA) in these plots. However, they're also associated with lowbush cranberry (VIBEDU), red osier dogwood (CORSTO), and bracted honey suckle (LONINV), shade tolerant shrubs that thrive in high light environments, but also persist under canopies in later successional stages (Abrahamson, 2022).
- Does decreased canopy cover and/or increased tree mortality lead to higher soil and microclimate temperatures?
- Is high tree cover associated with late-successional understory species?
- Are high volumesof recent deadwood associated with early-succesional understory species?
First of all, we can see that temperature has an orthogonal or slightly oppositely coorelated association with microclimate temperatures, indicating canopy cover has little to no effect on microclimate. Its a similar story with tree mortality: Temperatures are slightly corelated in figure 21, slightly negatively correlated in figure 22, and are correlated in figure 23, but the small length of the temperature vector indicates minor importance. Overall, I cant conclude that either tree canopies or recent deadwood has any significant effect on microclimatic hypothesis. My thought is that high understory covers in plots with high tree mortality are shading the microclimate sensors, resulting in lower temperatures for plots with less shading from the canopy.
Second, we can see that higher tree basal areas and canopy covers are associated with understory species that are shade tolerant, and late succesional. The association of species can depend on the type of canopy, with certain species like bog cranberry (VACVIT) and bishop's cap (MITNUD) preferring conifer forests, while a species like bunchberry (CORCAN) preferred deciduous canopies. Other species like wild sarsparilla (Picture 1, ARANUD), kidney-leaved violet (VIOREN), and wintergreen (PYRASA) occured readily under both canopy types. All the aforementioned species are shade tolerant, mid to late successional species that would decrease in importance after disturbance (Abrahamson, 2022).
Finally, we can also see that certain plants favour sites with higher volumes of recent deadwood: I found shade intolerant early succession plants such as fireweed (CHAANG), strawberry (FRAVIR), and birch seedlings (BETULA) in these plots. However, they're also associated with lowbush cranberry (VIBEDU), red osier dogwood (CORSTO), and bracted honey suckle (LONINV), shade tolerant shrubs that thrive in high light environments, but also persist under canopies in later successional stages (Abrahamson, 2022).
